The Anisian (Middle Triassic) Ammonoids of Nevada – An Integrated Approach to understand Morphological Change

1. Background
In Earth’s history, life struggled with five major mass extinctions. During these events, more than three-quarters of all species went extinct in a geologically short interval (Barnosky et al., 2011). The most severe extinction event took place at the Permian/Triassic boundary (e.g. Erwin, 1994; Benton, 1995), with 80–96 % of all marine species becoming extinct (Sahney and Benton, 2008).

The Mesozoic Marine Revolution (MMR)
Times after major mass extinctions always represent a resettlement of food webs including the predation pressure on individual groups of organisms. In his classic work Vermeij (1977) introduced the term “Mesozoic Marine Revolution (MMR)” which describes the full reorganization of predator-prey relationships after the Permian/Triassic extinction event. Vermeij hypothesized that the increase in predation, bioturbation, and competition led to the development of new defense strategies of the prey. This “arms race” led to changing morphologies and thus to new evolutionary pathways.
Ammonoids in Nevada
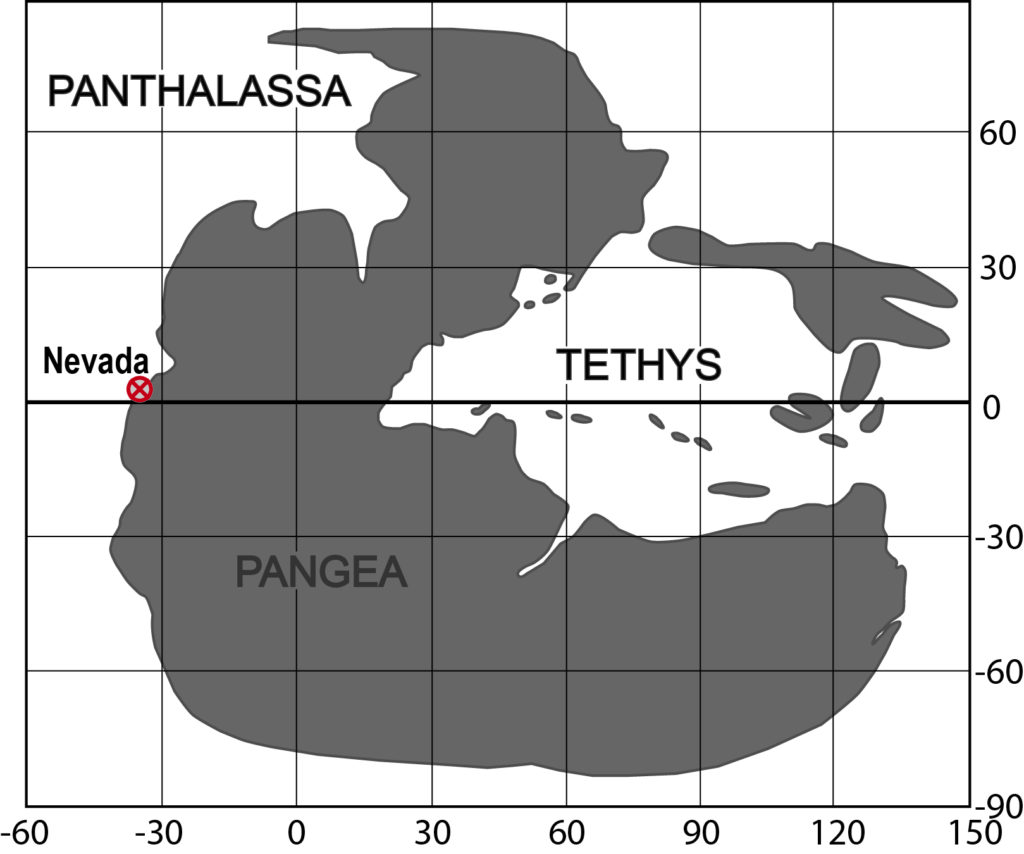
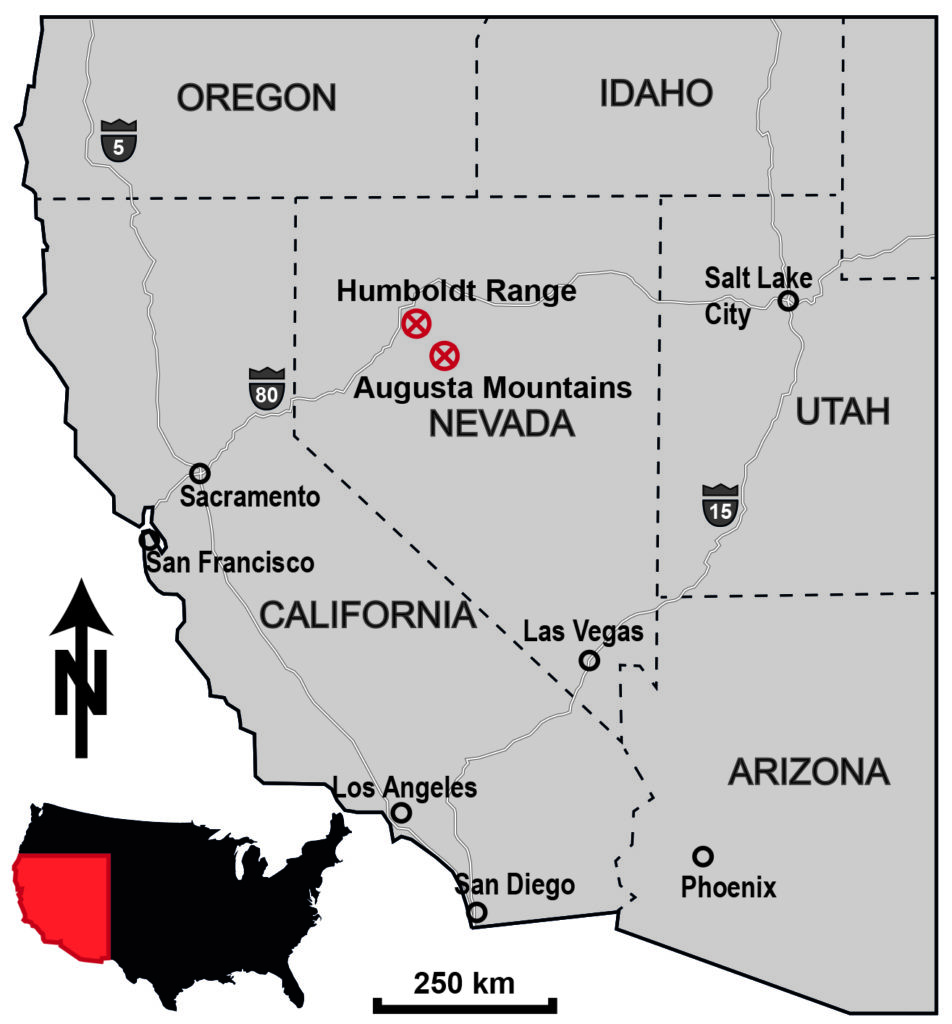
The Anisian (Middle Triassic) sediments that crop out in some mountain ranges in northwestern Nevada, USA, were deposited in a rather shallow part of the Panthalassic Ocean. Since it is particularly rich in fossils, the Fossil Hill Member (Prida and Favret Formation, Star Peak Group) is of particular interest to this study. This sequence consists of alternating layers of carbonates and marlstones. The two sequences in focus are located in the Humboldt Range and the Augusta Mountains.
The diverse Anisian ammonoid fauna of Nevada is well-known since the pioneer work of James Perin Smith (1914). A whole series of succeeding papers was revising this work and was refining the taxonomic approach (e.g. Silberling and Nichols, 1982; Monnet and Bucher, 2005). The taxonomic inventory is currently believed to comprise about 70 species (cf. Silberling and Nichols, 1982). However, by including fossil populations, the taxonomic concepts have been refined in recent years (for references see e.g. Monnet and Bucher, 2005). Simultaneously, a sophisticated zonal scheme has been developed.


Whereas fossils in the Augusta Mountains are mostly very well preserved, local intrusions in the Humboldt Range might cause slightly worse preservation (e.g. Lucas et al. 2007). Nevertheless, most fossils show distinct morphological features. Together with the very well documented, continuous stratigraphic sequence of the Fossil Hill Member, this forms a favorable basis for paleoecological studies.
2. Objectives
The broad goal of this project is to reveal interrelations between morphological change in ammonites and paleoenvironmental pressure. Since particularly predation pressure and its potential change is also directly expressed by the quantity and quality of shell damage, we plan to integrate paleopathology. Therefore, a large quantity of material is regarded as essential to unravel relationships.
The early phase of the Mesozoic Marine Revolution in the Anisian is expected to be a very sensitive interval with respect to predation pressure. This is hypothesized to be the case because this phase is characterized by changing environmental factors and a rise in the number of predatory groups. The latter became more and more important throughout the Mesozoic. The inclusion and analysis of both factors is necessary to judge the influence of environmental variations on morphological change and to discriminate it from other potential influence, such as shifts in the predator-prey relationship.
3. Methods
In order to ensure statistical relevance, this study is in need for a large number of specimens and is primarily designed quantitatively. The research aims are planned to be addressed by the following modules:
- Measuring of two lithostratigraphic sections, each one at the Fossil Hill (Humboldt Range) and one in the Muller Canyon (Augusta Mountains). Thereby samples for chemostratigraphy and microfacies analyses were obtained.
- Collecting of a large number of ammonoids bed-by-bed from selected stratigraphic levels.
- Processing of the collected raw material in the laboratory of the Geosciences Collection at the University of Bremen.
- Investigation of intraspecific variability and morphologic disparity in single faunal horizons. Thereby an inventory of morphological features using geometric and morphometric methods will be compiled.
- Identification of single groups using multivariate statistical software (PAST, R). This step also includes the photographic documentation of general morphologies. We will produce full images (lateral, dorsal, ventral) of around 80 specimens.
- Tracing the groups in the sections to unravel the expected morphological changes through time and to statistically compare single faunal horizons. Among other changes, heterochrony is very likely to encounter.
- Qualitative and quantitative analyses of ammonoid paleopathologies, also aided by SEM investigations. This is intended to measure different life styles of the fossil faunas.
- Investigation and interpretation of the carbonate microfacies and the chemostratigraphy (δ13C) of the measured sections.
4. Relevance of this study
Exploitation of ammonoids with respect to the “Mesozoic Marine Revolution” came into the focus of current research (Kerr and Kelley, 2015). The latter paper is concentrating on paleopathologies. In general very little is known about the predator-prey interaction with respect to ammonites as well as their morphological change. Additionally to modern, qualitative, individual case studies (e.g. Wright et al., 2014; Maddra, 2015), these topics recently move into top-ranking quantitative paleobiological research that allows an evidence-based scientific outcome (Takeda and Tanabe, 2015). Therefore the investigation of morphological variation as a consequence of paleoenvironmental perturbations, ontogeny, evolution, and developmental instability is a seminal field, which is addressed in various recent publications (e.g. Klug et al., 2016; Lehmann et al., 2016; Owniak, 2012; Patzkowsky and Holland, 2012; Schlüter, 2016). Despite their importance, most studies on this field are pioneering work, rarely addressed in invertebrate paleontology. An obstacle hindering the investigation of morphological change in paleobiogical studies is the rare occurrence of suitable outcrops. Each individual section investigated should be continuous and should contain a large number of well-preserved specimens. Furthermore, the preservation modes of fossils investigated need to be similar comparing results of different localities.
Keeping this in mind, the working area in NW Nevada, focused on here, offers excellent conditions to study morphological change in paleobiology. In contrast to many other localities in the world, the study sites herein are continuous successions, exhibiting large numbers of specimens. Therefore, a high temporal and spatial resolution through bed-by-bed collecting can be achieved. This project aims to exploit the promising test range of the Triassic in NW Nevada. The observation of the presumed taxonomic and morphological changes through time will help to understand evolutionary patterns.
Integrating of chemostratigraphy, namely δ13C stable isotopes, will tighten the stratigraphical scheme of the region. According to Lucas et al. (2007) there is no δ13C record published for the Triassic of Nevada. As carbon isotopes are a valuable tool for correlation of individual sections, this study also provides important basic data for the stratigraphy of this area and period.

References:
- Barnosky, A.D., Matzke, N., Tomiya, S., Wogan, G.O.U., Swartz, B., Quental, T.B., Marshall, C., McGuire, J.L., Lindsey, E.L., Maguire, K.C., Mersey, B., Ferrer, E.A., 2011. Has the Earth/’s sixth mass extinction already arrived? Nature 471, 51-57.
- Benton, M.J., 1995. Diversification and Extinction in the History of Life. Science 268, 52-58.
- Erwin, D.H., 1994. The Permo-Triassic extinction. Nature 367, 231-236.
- Klug, C., De Baets, K., Korn, D., 2016. Exploring the limits of morphospace: Ontogeny and ecology of late Viséan ammonoids from the Tafilalt, Morocco. Acta Palaeontologica Polonica 61, 1-14.
- Lehmann, J., von Bargen, D., Engelke, J., Claßen, J., 2016. Morphological variability in response to palaeoenvironmental change – a case study on Cretaceous ammonites. Lethaia 49, 73-86.
- Maddra, R., 2015. Bitten ammonites from the upper Lias Group (Lower Jurassic) of Saltwick Bay, Whitby, North Yorkshire, UK. Proceedings of the Yorkshire Geological Society 60, 153.
- Monnet, C., Bucher, H., 2005. New Middle and Late Anisian (Middle Triassic) ammonoid faunas from Northwestern Nevada (USA): Taxonomy and biochronology. Fossils and Strata 52, 1-121.
- Owniak, E., 2012. The perisphinctid genus Prososphinctes Schindewolf (Ammonoidea, subfamily Prososphinctinae nov.): an indicator of palaeoecological changes in the Early Oxfordian Submediterranean sea of southern Poland. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 264, 117-179.
- Patzkowsky, M.E., Holland, S.M., 2012. Stratigraphic Paleobiology. Understanding the distribution of fossil taxa in time and space, The University oc Chicago Press, Chicago, p. 256.
- Péron, S., Bourquin, S., Fluteau, F., Guillocheau, F., 2005. Paleoenvironment reconstructions and climate simulations of the Early Triassic: Impact of the water and sediment supply on the preservation of fluvial systems. Geodinamica Acta 18, 431-446.
- Sahney, S., Benton, M.J., 2008. Recovery from the Most Profound Mass Extinction of All Time. Proceedings: Biological Sciences 275, 759-765.
- Schlüter, N., 2016. Ecophenotypic variation and developmental instability in the Late Cretaceous echinoid Micraster brevis (Irregularia; Spatangoida). PLoS ONE 2016, 1-26.
- Silberling, N.J., Nichols, D.J., 1982. Middle Triassic molluscan fossils of biostratigrahic significance from the Humboldt Range, northwestern Nevada. U.S. Geological Survey Professional Paper.
- Smith, J.P., 1914. The Middle Triassic marine invertebrate faunas of North America, Professional Papers U.S. Geological Survey 83. Washington, p. 254.
- Takeda, Y., Tanabe, K., 2015. Low durophagous predation on Early Toarcian (Early Jurassic) ammonoids in the northwestern Panthalassa shelf basin. Acta Palaeontologica Polonica 60, 781-794.
- Vermeij, G.J., 1977. The Mesozoic Marine Revolution: Evidence from snails, predators and grazers. Paleobiology 3, 245-258.
- Wright, J.K., Bassett-Butt, L., Collinson, M., 2014. Fatally bitten ammonites from the Lower Calcareous Grit Formation (Upper Jurassic) of NE Yorkshire, UK. Proceedings of the Yorkshire Geological Society 60, 1.
Within the project “Science for the living room” the research was presented in a lecture (in german).
